Rubrique : AnimauxAuteur : vonvonNiveau : Confirmé
INTRODUCTION :
On trouve de nos jours de nouvelles espèces de coraux disponibles sur le marché aquariophile dont beaucoup sont agressives. Une attention toute particulière doit être apportée à réduire l'agression des coraux entre eux. En effet, si par le passé maintenir un corail relevait du défi, les avancées faites dans le domaine de l’aquariophilie récifale (tant technologiques que des nouvelles méthodes de maintenance) font que les coraux hébergés dans nos bacs deviennent de véritables colonies à la croissance rapide et à la taille imposante. À mesure que la taille de ces coraux augmente ajouté à la petitesse de l’habitat que nous leur offrons, la proximité des uns aux autres a pour conséquence la manifestation de leur nature agressive. Ainsi, alors que leur agressivité était à peine observable et pas un problème quand ils étaient encore de petite taille, leur effet sur les coraux voisins devient nettement apparent quand ils atteignent une taille conséquente.
Et oui, la nature est bien faite, chaque espèce animale, végétale ou microbienne (et humaine…) n’a qu’un seul but : transmettre les gênes. Pour cela chacun a sa parade pour préserver son territoire de prédilection et conserver des conditions idéales à sa prolifération. Même les coraux les plus passifs doivent pouvoir tenir leur espace vital, sinon ils seront recouverts et éliminés par d'autres espèces. A noter que certains coraux sont même urticants pour l'Homme et pouvant donc provoquer des allergies !
Nous allons donc nous intéresser aux différentes techniques de combat que se livrent les Coraux, en connaître les causes et remèdes. Cela implique une observation et connaissance particulière des hôtes de nos bacs, car si différentes espèces de coraux se livrent des batailles chimiques, elles ne le font pas forcément de jour et préfèrent chasser la nuit, sortant des polypes qui peuvent entrer en contact avec leurs voisins. Cela sans compter le déploiement de l’animal, par exemple l’Euphyllia spp ne mesurant pas le même diamètre en pleine journée que la nuit.
Pour commencer une petite définition des Cnidaires :
Les Cnidaires (appelés autrefois Cœlentérés) sont présents sur terre depuis plus de 570 millions d'années et regroupent plus de 10.000 espèces dont une centaine sont dangereuses pour l'Homme. Ils représentent un embranchement d'invertébrés représentés par les Hydres, Méduses (forme libre), les Anémones (polype fixé) et les Coraux (squelette calcaire). Ils sont carnivores et caractérisés par une cavité digestive (cœlentéron) ouverte par une bouche et entourée de tentacules. On distingue 4 classes : Hydrozoa, Scyphozoa, Cubozoa et Anthozoa. Leur appareil venimeux est constitué par le cnidocyte, cellule sécrétrice et sensorielle qui renferme une capsule, le cnida (nématocyste ou spirocyste ou ptychocyste) d'une taille généralement inférieure à la centaine de microns. Il contient du venin et un tube enroulé en hélice. Les venins sont des amalgames de haut poids moléculaire, constitués de toxines, d'amines actives sur les vaisseaux sanguins et d'enzymes détruisant les protéines. Sous l'effet d'un stimulus mécanique ou chimique, le tube jaillit comme un ressort en injectant le venin. Il peut exister plus de 100 000 nématocyste/cm sur un tentacule de méduse.

Schéma du nématocyste. A gauche, avant la décharge, à droite, après la décharge. Cc : contenu capsulaire, Ep : épines, H : hampe. Op : opercule pn : paroi, Ter : tube terminal (d'après Weill, 1934)
DIFFERENTS TYPES D’AGRESSION DU CORAIL :
Croissance rapide :
Le taux de croissance des coraux durs diffèrent selon les espèces, certaines ayant une croissance beaucoup plus rapide dans les eaux peu profondes que d'autres. C'est un avantage qui permet à ces coraux de coloniser rapidement de nouveaux secteurs. Une vitesse de croissance rapide permet également à ces espèces de réaliser la dominance sur d'autres par « overtopping », traduisez « pousser par dessus » réduisant de ce fait la quantité de lumière et de circulation d'eau qu’elles peuvent recevoir (Huston, 1985). Ce mécanisme a été proposé comme explication de la dominance des coraux tels que le Pocillopora dans l’océan Pacifique et l'Acropora dans l'océan Atlantique (Huston, 1985), à noter que le Millepora a aussi cette faculté. Ce mécanisme est d'importance limitée en aquarium dû à la faible proportion de coraux durs. Cependant, l'amateur ne devrait pas négliger les effets d' « overtopping » causés par de grandes Anémones, Coraux (mous ou durs) ou Macroalgae, ces dernières provoquant le blanchiement du corail attaqué. Ceci peut potentiellement se produire dans un aquarium et on devrait toujours s'assurer qu'un spécimen reçoit suffisamment de lumière et une bonne circulation d'eau.
Il est préférable d’allouer un espace spécifique pour les coraux à croissance rapide et de placer les autres espèces en fonction.
-> Microdictyon (Macroalgae) recouvrant un Montastrea annularis
-> Acropora 
-> Millepora complenata colonisant un Montastrea annularis (photo du haut) et une gorgone (photo du bas)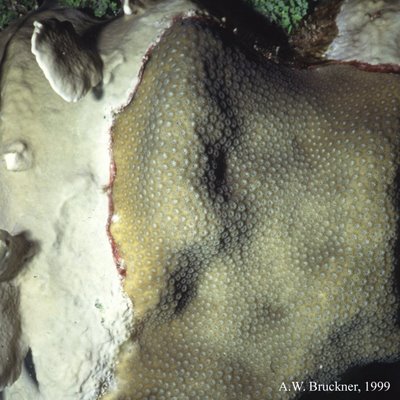

-> Pocillopora
Filaments mésentériques :
Plusieurs espèces de coraux durs peuvent produire des filaments de Mesenterial (également nommés filaments mésentériques) à partir de leur estomac. Les coraux des genres Favia, Favites, Scolymia, Pavona, Euphyllia, Cynarina et certains Acroporas ont tous cette capacité (Chadwich, 1987).
Les filaments mésentériques peuvent être employés tout à fait efficacement en tant que structures agressives, et dévorer d'autres polypes de corail par un processus semblable à la digestion.
Quand deux coraux durs entrent en contact (différentes espèces ou les mêmes), l’agresseur expulse des filaments mésentériques par la cavité de sa bouche ou de ses tissus sur la surface de l'autre, digérant ainsi les tissus de son voisin. Ceci a comme conséquence une zone de squelette nue qui peut alors être envahie (Sebens et Miles, 1988). Cette zone peut être envahie par le corail attaquant ou permettre de créer une « zone tampon » entre les deux espèces (Huston, 1985).
-> Déploiement de filaments mésentériques d'un Hydnophora rigida sur un Stylophora pistillata :

Quelques coraux, comme l’Euphyllia, ont même la capacité de produire des tentacules répulsives (voir plus bas) et des filaments mésentériques, leur permettant ainsi de livrer bataille sur plusieurs fronts (Wallace, 1984).
-> Favites
-> Cynarina
-> Scolymia
-> Favia
-> Acropora millepora

-> Euphyllia

Acrorhagi :
Ce sont des structures spécialisées qui ont été identifiées la première fois sur des anémones d’eau froide de la famille des Actiniidae. Elles se composent de sacs gonflés de cnidoblastes, caractère propre des cnidaires, qui sont des cellules urticantes spécialisées (appelé aussi nématoblaste), appareil venimeux servant à la défense et à la capture des proies ainsi qu'à l'adhérence au substrat. Les cnidoblastes sont nombreux au niveau des tentacules et ne servent qu'une seule fois, lorsque le cnidocyste s'est ouvert et que le venin a été évacué à l'extérieur, le cnidoblaste meurt et est remplacé par un autre, issu d'une cellule interstitielle.
-> Acrorhagi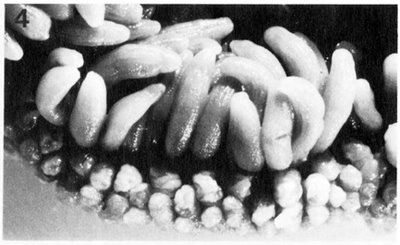
-> Cnidoblaste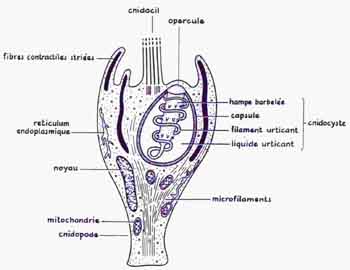

Le cnidocyste est une vacuole (organite cellulaire) dont la longueur varie de 10 à 100 µm, contenant un filament urticant dévaginable. On distingue différents types parmis lesquels les spirocystes, les homotriches (= holotriches), les ptichocystes et les nématocystes.
-> Cnidocyste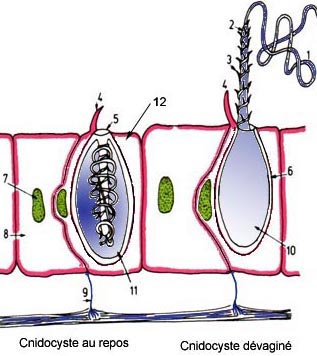
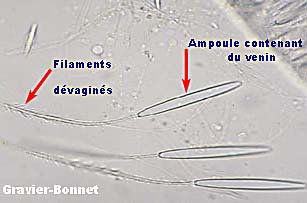
On parle aussi d’acrosphère qui est l’extrémité renflée et chargée de cnidoblastes d'une tentacule. Quand elles sont en contact avec une autre anémone elles laissent une couche du tissu qui a comme conséquence la mort localisée du tissu de l'intrus (Sebens et Miles, 1988).
-> Polype de Pocillopora agrandi 30X montrant les Acrosphères blancs et ronds à l'extrémité de chaque tentacule : 
Outre les anémones possédant des cnidoblastes et ayant la faculté de se déplacer, et donc de brûler les coraux par contact, on retrouve aussi des cellules urticantes chez les espèces telles que :
-> Discosomas

-> Ricordéas
-> Zoanthus
-> Parazoanthus
-> Palythoa
Dans la même lignée il existe aussi les Aconties, prolongements filamenteux de la base du lobe médian des cloisons gastriques de certaines anémones (excepté les Actiniidae). Ces filaments, contenant des enzymes digestives, peuvent être rejetés hors de la cavité gastrique au travers de la colonne, voire du disque oral, pour la défense ou l'agression.
Tentacules répulsives :
Les tentacules répulsives sont des tentacules spécialisées dans la protection du territoire, soit par la défense soit par l’attaque. Elles apparaissent sur des polypes après plusieurs semaines de contact avec d'autres colonies (Hidaka et Yamazato, 1984 ; Sebens et Miles, 1988), mais certains coraux comme le Galaxea semblerait en avoir en permanence. Ces tentacules sont habituellement beaucoup plus longues et ont beaucoup plus de cellules urticantes (nématocystes) que la normale, elles peuvent attaquer un corail entré en concurrence et littéralement le « brûler ». En conséquence leur fonction a changé, passant de celle d'une d'alimentation à celle de la défense ou de l'agression. Des polypes ovales de certains coraux tels que Goniopora, peuvent également être employés en tant que « polypes répulsives » dans un but agressif. Bien que la production des tentacules répulsives soit habituellement associée aux coraux durs, une étude récente a prouvé qu'ils peuvent également se développer chez les coraux mous (sous-classe des Octocorallia) comme l’Erythropodium caribaeorum (Sebens et Miles, 1988). Ces tentacules répulsives sont réparties sur le contour de la colonie et chargées de nématocystes prêts à être expulsées au contact d’un autre corail (Hidaka et Yamazato, 1984).
-> Erythropodium caribaeorum colonisant un Siderastrea siderea (photo du haut) et un Montastrea annularis (photo du bas)

Ates (1989) fournit une liste des coraux durs agressifs, dont certains sont souvent maintenus dans des aquariums tels que le corail bulle (Plerogyra sinuosa), les espèces d’Euphyllidae (Euphyllia ancora et divisa), de Favia, le Catalaphyllia jardinei, les espèces de Favites et les espèces de Galaxea. Thomason et Brown (1986) ont constaté qu'il y avait un rapport direct entre la compétence agressive et le nombre de nématocystes par polype et/ou le filament mésentérique. Une chose intéressante est constatée, c'est le nombre de nématocystes par structure qui est important et non pas la taille ou le nombre de polypes et de filaments mésentériques. Combinant leurs résultats avec ceux de Sheppard (1979), divers coraux de l’Indo-Pacifiques peuvent être placés dans les catégories agressives suivantes :
-> Plerogyra sinuosa
-> Catalaphyllia jardinei
-> Euphyllia ancora
-> Fungia
-> Goniopora (nématocystes concentrés dans les polypes et non pas dans les filaments mésentériques)

-> Galaxea
-> certaines espèces d'Acropora
-> Lobophyllia
-> Montipora
-> Porites
-> Platygyra daedalea
-> Trachyphyllia geoffroyi
-> Goniastrea favulus
-> Caulastrea
Dans Caribbean, Lang (1973) a classifié l'Isophyllia sinuosa en tant que très agressif, le Montastrea annularis en tant qu'espèce modérément agressive et le Porites comme faiblement agressives. La dominance principale du Montastrea est due entièrement à l'action de ses filaments mésentérique. Cependant, Bak et autres (1982) ont constaté qu'à mesure que l’espace entre deux espèces se réduisait, le corail subalterne pouvait développer des tentacules répulsives et renverser l'interaction en sa faveur.
-> Isophyllia sinuosa
-> Montastrea
Composés toxiques :
Sur Terre, sous les eaux douces ou marines, des animaux produisent des métabolites secondaires qui peuvent agir en tant que signaux chimiques sur leurs concurrents (Maida et. Al, 1995). Ce sont ces signaux chimiques qui sont les armes mortelles dans la guerre pour l'espace parmi les organisations sessile. Ces signaux peuvent être inhibiteurs ou stimulatoires. C'est par les produits chimiques inhibiteurs que les premiers « résidants » peuvent rendre l'environnement physique plus ou moins favorable pour des concurrents voulant s’approprier cet espace (Breitburg, 1985). Dans des conditions de forte concurrence au sein d'un récif corallien, c'est un avantage d’avoir des toxines qui sont efficaces contre les concurrents sessiles potentiels. De telles toxines sont connues comme allélochimiques, elles empêchent la capacité d'une autre colonie à partager le même habitat (Fearon et Cameron, 1997). Quelques exemples d’organisations marines fortement toxiques : l’étoile de mer Acanthaster, le poisson pierre Synanceia, le mollusque Conus geographus, et le crabe Zosimus aeneus. Généralement, les défenses chimiques sont trouvées dans les organisations n’ayant pas d'autres moyens de défense tels que la capacité de s'échapper.
En 1971, Robert Whittaker et Paul Feeny de l’Université de Cornell, proposent de nommer substances allélochimiques les métabolites produits par un individu d’une espèce et capables d’affecter la croissance, la santé, la biologie ou le comportement d’une autre espèce. Par la suite, ces chercheurs distinguent deux types de substances allélochimiques : les allomones qui procurent un avantage à l’organisme qui les émet (substances défensives) et les kairomones
qui procurent un avantage à l’organisme qui les perçoit.
Les substances, phéromones, allomones et kairomones, impliquées dans ces phénomènes de communication chimique ont été qualifiées de métabolites secondaires, par opposition aux métabolites primaires. Les métabolites primaires sont synthétisés normalement par l’organisme pour sa croissance et sa reproduction ; ils sont communs à tous les organismes vivants, ils traduisent l'uniformité du monde vivant. Les métabolites secondaires sont des produits dérivant du métabolisme général et ne jouent apparemment aucun rôle vital ; ils sont propres à chaque espèce, ils sont l'expression de la diversité du monde vivant.
Définition de allélopathie : Influence réciproque des plantes (autres que des micro-organismes), les unes sur les autres, résultant des produits de leurs métabolismes. Cette compétition nocive est exercée entre des plantes d'espèces différentes, par l'intermédiaire de substances toxiques excrétées par les racines, les feuilles ou la litière.
Le biotope marin est considéré comme le plus riche des différents habitats du globe mais également comme le moins bien connu des scientifiques. C'est aussi le berceau prébiotique de notre planète (3 milliards d'années d'évolution !) ; ceci a pour conséquence une diversité d'espèces, de systèmes d'organisation, de formes et de solutions adaptatives. Cette biodiversité est la source d’une formidable chimiodiversité.
En milieu marin, les systèmes de communication chimique constituent un élément indispensable dans l'établissement des relations intra et interspécifiques. Dans l'eau, la signalisation visuelle perd une grande partie de son intérêt ; de très nombreux invertébrés ne possèdent d'ailleurs pas d'organe de vision.
Depuis plus de cent millions d'années, le milieu marin vit donc une course aux armes chimiques, il est le siège de luttes incessantes entre proies et prédateurs ou entre colonisateurs et colonisés. La fonction vectrice de l'eau où s'exercent diffusion et dispersion avec une grande facilité, favorise l'établissement de barrières chimiques dressées à l'aide de substances organiques. Ces substances peuvent réduire la valeur nutritive des tissus, être désagréables au goût (antipalatabilité), toxiques ou antifouling.
Le milieu marin est très riche en exemple de substances allélochimiques de toutes sortes, des plus simples aux plus complexes ; la même molécule pouvant jouer le rôle d’allomone ou de kairomone. La capacité à synthétiser des médiateurs chimiques impliqués dans la défense et la compétition interspécifique est plus particulièrement développée chez les animaux sessiles c’est-à-dire vivant toujours fixés au substrat comme le sont les végétaux. De nombreux invertébrés marins (Coraux, Gorgones, Eponges, Ascidies, etc.) ont en effet un mode de vie fixé et sont d'un point de vue écologique identiques aux végétaux ; ils sont immobiles et photosynthétiques pour beaucoup par le biais des microorganismes symbiotiques qu'ils renferment. Incapables dans la plupart des cas de réponses comportementales à une agression, ils sont pourvus de protection physique ou chimique ou de caractéristiques morphologiques pour se protéger des prédateurs, compétiteurs ou autres organismes pathogènes.
Chez les Coraux :
Au commencement… :
Le cycle de vie des Coraux inclut une phase larvaire planctonique qui peut durer de quelques minutes à quelques mois (Pawlik, 1992). Cela signifie que quelques larves peuvent voyager pour seulement quelques mètres tandis que d'autres peuvent voyager sur des centaines de kilomètres. Il y a deux genres de larves :
- Les larves Planctotrophiques s’alimentant et ayant différents stades de développement sur une longue période.
- Les larves Lecithotrophiques ne s'alimentant pas et ayant une période de développement courte (Pawlik, 1992).
En raison de la concurrence intense pour l'espace dans des écosystèmes marins, l'endroit dans lequel les larves choisissent de s’installer est extrêmement important. La difficulté des larves à localiser la nourriture ou d'autres ressources a comme conséquence la mort, et les larves doivent éviter de s'installer dans des secteurs hostiles et fortement concurrentiels (Young et Chia, 1981). Le procédé de règlement des larves planctoniques est influencé par beaucoup de facteurs environnementaux. La plupart des larves se sont avérées s’installer au premier contact avec le substrat (63%), et 6.7% des larves ont fait dix contacts ou plus avant de s’installer. Certains des facteurs physiques influençant le règlement larvaire peuvent inclure la lumière, la pesanteur, la pression hydrostatique, la température, et la salinité (Pawlik, 1992). Les propriétés du substrat comme la texture ou la température peuvent également être impliquées (Pawlik, 1992).
Évidence des toxines allélochimiques :
Les produits chimiques Allélopathiques sont produits par une organisation pour inhiber la capacité d'une autre organisation à partager le même habitat. Ces toxines sont efficaces contre le concurrent sessile potentiel car hostiles aux organisations autre que celles qui les produisent. L'utilisation des produits chimiques pour empêcher la croissance des espèces des unes par les autres s'appelle l'allélopathie, terme commun aux plantes terrestres, dans notre cas cela s'appelle l'amensalisme.
Définition de amensalisme : relation entre deux espèces d'un même biotope se traduisant par l'inhibition du développement de l'une d'entre elles.
- les coraux durs :
Les principaux bâtisseurs de récif, ou coraux hermatypiques sont les coraux de l’ordre des scléractinaires (Fearon et Cameron, 1997). Il a été mis en évidence le dégagement de toxines commun à beaucoup d'invertébrés sessile coloniaux. Ces produits chimiques toxiques appelés larvotoxines ont été trouvées chez les coraux hermatypiques de la famille des Faviidae (ex : Platygyra daedalea, Favia matthai et Goniastrea favulus), Fungiidae (ex : Fungia fungites), Agariciidae (ex : Pavona decussata), Poritiidae, et Dendrophylliidae (Fearon et Cameron, 1997). Parmi les Scléractinaires, 71% de 58 espèces examinées étaient toxiques aux souris, 86% de 57 espèces examinées ont une activité hémolytique, 45% des 55 espèces examinées possèdent une activité antimicrobienne, et 9% de 45 espèces étaient ichtyotoxique, c'est-à-dire toxique pour les poissons (Gunthorpe et Cameron, 1990a). Le rôle de ces larvotoxines peut être d'affecter le règlement et la prolifération des concurrents potentiels (Fearon et Cameron, 1997). Ils ont observé que les Coraux réagissent aux toxines avec une contraction soutenue des polypes, protection accrue de mucus, perdent de la coloration de tissu, et de la perte de tissu squelettique. Bien que ces coraux durs contiennent clairement des toxines, la question demeure si ces toxines sont libérées dans leur environnement normal. Comme précisés par Jackson et Buss (1975), des toxines peuvent juste être stockées à l'intérieur de l'organisation mais non dégagées dans l'environnement.
-> Fungia fungites (Fungiidae)
->Heteropsammia cochlea (Dendrophylliidae)
-> Turbinaria (Dendrophylliidae)
-> Pavona cactus (Agariciidae)
- les coraux mous :
Outre les coraux hermatypiques de l’ordre des Scléractinaires il y a les Alcyonaires, terme général regroupant les stolonifères, les coraux mous et les gorgones à l'exclusion des pennatules. Il est bien connu que beaucoup de coraux mous contiennent de nombreux composés toxiques tels que des terpénoïdes (exemple le Clavularia, Pachyclavularia viridis, Xenia et Zoanthus), et la sarcophine (exemple le Sarcophyton glaucum). Ces produits chimiques se sont avérés très efficaces dans le découragement des prédateurs mais suggère aussi qu'ils soient utilisés comme mécanisme pour maintenir un espace vital (Sheppard, 1979). Coll et autres (1982) ont isolé des terpènes toxiques de l'eau entourant plusieurs coraux mous tel le Sinularia.
Définition de terpène : hydrocarbure des huiles essentielles volatiles, ayant une forte odeur, synthétisé par les coraux mous pour éloigner les prédateurs.
-> Sarcophyton glaucum
-> Pachyclavularia viridis
-> Clavularia
-> Sinularia flexibilis
-> Lobophytum pauciflorum
-> Xénia

Interaction entre les Alcyonaires et les Acléractinaires :
Expériences en milieu clos :
Dans diverses expériences utilisant des greffes de Lobophytum pauciflorum, de Sinularia pavida et des espèces de Xenia, Sammarco et autres ont relevés des effets sur le Pavona cactus et le Pseudosuberites andrewsi (Porites) quand les coraux mous ont été placés près des colonies ou en contact direct avec elles. Ils ont conclu que les effets des coraux mous sur les coraux durs étaient entièrement spécifiques à l'espèce. Dans certains cas, une distance de 30 centimètres entre les colonies a eu comme conséquence la croissance stoppée puis la mort du corail dur, alors que d’autres espèces de coraux mous l’endommageaient seulement malgré un contact direct. Par exemple, le Pseudosuberites andrewsi était le corail dur le plus sensible, réagissant à deux des trois coraux mous utilisés, même à une distance de 10 centimètres. Cependant, un corail mou qui peut endommager par le contact ne peut pas nécessairement le faire en déchargeant des composés dans l'eau. Par exemple, l’espèce de Xenia a endommagé les deux coraux durs examinés seulement au contact. D'autre part, si un corail mou (par exemple Lobophytum pauciflorum) peut endommager un concurrent à distance, il peut également le faire par contact (Sammarco et autres, 1983). Par l'utilisation des composés toxiques, les coraux mous peuvent concurrencer les coraux durs pour l'espace en arrêtant leur croissance voire même en tuant toute ou partie d'une colonie et repousser sur ce nouvel espace. Cependant, les coraux durs ne sont pas sans leurs propres défenses et des dommages sérieux peuvent aussi se produire sur les coraux mous.
Expériences en milieu naturel :
Une grande partie de la recherche faite avec le Corail dans son environnement normal implique les coraux mous fortement toxiques et leurs effets sur les coraux durs. Sammarco et autres (1983) ont effectué leurs expériences sur des Alcyonaires et Scléractinaires cohabitant à proximité étroite. D’après leurs résultats, dans 17% des cas ils ont observés une nécrose des tissus des Scléractinaires situés à proximité immédiate du corail mou. Dans 52 % des cas observés, il a été relevé des effets mortels secondaires allant de l'inhibition à l'arrêt de la croissance du corail dur. Le Lobophytum pauciflorum s'est avéré causer la nécrose des tissus du Pseudosuberites andrewsi par contact et à distance, le même cas s’est produit avec le Lobophytum sur le Pavona mais aucun effet n’a été constaté sans contact. Le Xenia a causé la nécrose des tissus du corail dur par contact, mais n'a eu aucun effet autrement, même chose pour le Sinularia pavida avec le Pseudosuberites andrewsi. Maida et autres (1995a, 1995b) ont constaté que le Sinularia flexibilis, a eu un effet inhibiteur directionnel sur les coraux durs, plus particulièrement en bas du courant où se trouve le corail mou. Les coraux durs examinés n'ont pas eu cet effet inhibiteur. Cependant il est difficile de préciser si les larves des Scléractinaires ont senti les composants allélochimiques du Sinularia flexibilis et évités le secteur, si elles se sont tout d’abord posée sur le substrat avant de le quitter, ou bien de s’y installer avant de mourir et sécréter un squelette de carbonate de calcium. Maida et autres (1995a, et 1995b) sont venus à ces quatre conclusions suite à leurs recherches :
- Les coraux mous (Alcyonaires) sont des concurrents efficaces pour l'espace vis-à-vis des coraux durs (Scléractinaires) en causant une mortalité localisée de ces derniers.
– La mortalité peut être causée par le contact direct ou à distance par relarguage dans l'eau d’exsudats allélopathiques.
- Les capacités concurrentielles changent d'une façon spécifique à l'espèce.
- La susceptibilité des Scléractinaires change également d'une façon spécifique à l'espèce.
En parallèle, des recherches ont été faites par Coll et autres (1982a) et ont isolé du flexibilide et du dihydroflexibilide, aussi bien que les terpènes, de la sarcophine et sarcophytoxide de l'eau de mer autour des deux coraux Sinularia flexibilis et Sarcophyton crassocaule, démontrant que ces composés sont déchargés dans l'eau. Avec ces terpènes, des lipides et stérols ont été également isolés dans l'eau de mer entourant les coraux mous. Aucun de ces derniers composants chimiques n'est très hydrosoluble, ainsi on a présumé que cette capacité de relarguage dans l'eau peut être due au dégagement de mucus par la colonie (Coll et autres. 1982a).
Cependant, beaucoup de preuves suscitent l'idée que ces composés toxiques dans les coraux mous servent de défense et permettent de plus à concurrencer l'espace (Bakus, 1986). Basé sur des analogies avec les systèmes terrestres, il existerait une variation de la qualité et de la quantité de composants allélochimiques en réponse aux changements de l'environnement ou des pressions sélectives (Feeny, 1976). Bakus (1981) a constaté qu'une incidence plus élevée de la toxicité se produit chez les espèces où la prédation de poissons et le pâturage sont plus intenses, et que les éponges de récifs coralliens, les coraux mous, ou les ascidies qui manquent de composants allélochimiques ont un corps durci ou sécrète des quantités énormes de mucus (Bakus et autres. 1986). Ces défenses mécaniques peuvent supplanter le besoin de défenses chimiques contre la prédation.
Conclusion :
Si un certain type de corail contient des composants chimiques toxiques, cela ne signifie pas nécessairement qu'il les décharge dans l’eau ou qu’ils soient impliqués dans l'allélopathie. Même si les exsudats ne sont pas mortels, ils peuvent tout de même donner un avantage à la colonie établie en limitant la croissance ou la physiologie des coraux voisins (Fearon et Cameron, 1997). La capacité des larves à détecter et éviter les exsudats qui peuvent plus tard être mortels après exposition chronique et à long terme est un avantage (Fearon et Cameron, 1997). Cette bataille incessante pour l’espace, impliquant des composants allélochimiques, peut être établi dans le mécanisme pour maintenir la diversité dans des écosystèmes marins, et que donc toutes ces interactions sont au final d’une valeur inestimable quant au fonctionnement et à l’établissement de ces écosystèmes si fragiles.
FIN DE LA PREMIERE PARTIE
cliquez ici pour la deuxième partie
Origine des textes :
« La communication chimique dans le monde vivant »
Bernard BANAIGS Inserm, Université de Perpignan
« Les envenimations marines »
Dr Jean-Yves BERNEY Hôpital Cantonal Universitaire de Genève
“The Role of Allelopathic Chemicals in the Spatial Patterning of Benthic Communities”
Ali LINDBERG Biology Department Colorado State University Fort Collins, CO, 80523
“Coral Compatibility”
J. Charles DELBEEK B.Ed M.Sc. Sept/Oct. 1990 Aquarium Fish Magazine
“Coral placement and aggression »
GREGG and SALTMAN - April 27, 1997
Remerciements :
jlc, 27-L, Coco_bel_oeil, joann7, fredb, typedu94, Systemc, Coyote et geka pour leurs superbes photos, elgoog pour la traduction (souvent difficile) et tout les chercheurs, biologistes passionnés et récifalistes attentionnés qui nous font découvrir un peu plus chaque jour la fabuleuse vie sous-marine, berceau de la vie...
Liens :
http://www.uea.ac.uk/env/all/faculty/grant/Interactions%20on%20hard%20substrates.htm
http://www.reef-guardian.com/la-bataille-chimique-pour-lespace-898-imprimer.html
http://www.colostate.edu/Depts/Entomology/courses/en570/papers_1998/lindberg.htm
http://www2.hawaii.edu/~delbeek/reefaq6.html
http://calypsoplongee.free.fr/gbio.htm
http://scaphinfo.free.fr/bio/envenimation.html
http://www.petsforum.com/personal/trevor-jones/coralfeeding.html
http://www.unep-wcmc.org/marine/coraldis/cd/overgrowth.htm
http://www.univ-perp.fr/perspectives/IMG/pdf/communication_chimique.pdf#search=%22Communication%20chimique%20dans%20le%20mode%20vivant%20de%20BERNARD%20BANAIGS%22
























































 Les articles
Les articles

