Photo : jlc  |
Les coraux mous : ce sont des Alcyonacea (sous-classe des Octocorallaires, classe des Anthozoaires, embranchement des Cnidaires).
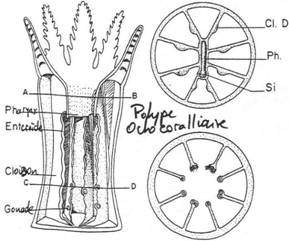 La circulation de l'eau :
La circulation de l'eau :La circulation de l’eau au sein des colonies coralliennes joue un rôle fondamental dans le métabolisme du corail : elle facilite la nutrition, permet les échanges gazeux et favorise les échanges de matière avec le milieu extérieur.
La circulation de l’eau chez les Octocoralliaires est assurée grâce aux cils, ces derniers sont situés à l’extrémité ventrale du pharynx (chaque polype possède un pharynx) et forment une gouttière. Cette gouttière bordée de cellules ciliées est destinée à assurer la circulation de l’eau dans la cavité gastrique et pourrait participer à l’ingestion de particules alimentaires. Les cils provoquent des mouvements à l’origine d’un déplacement d’eau appelé « courant ciliaire ».
 La nutrition :
La nutrition :La nutrition est assurée pour la presque totalité des espèces par de petits organismes planctoniques, cependant certains Octocoralliaires peuvent absorber nutriments et minéraux contenus dans l’eau ainsi que du mucus porteur de bactéries et de matières organiques.
Chez les coraux mous, la majeure partie du plancton absorbé est constituée par le phytoplancton (représenté par de nombreuses espèces d’algues comme les Diatomées et les Dinoflagellés). Concernant le zooplancton (plancton animal représenté par de petits crustacés et leurs larves ainsi que par les larves de mollusques, d’annélides et de poissons), les coraux mous ont une prédilection pour le plancton peu actif et présentent une incapacité à piéger des proies plus mobiles ; cela est très probablement dû à un manque d’efficacité des cnidoblastes (Sprung et Delbeeck, 1999).
 La capture est assurée par le courant inhalant du siphonoglyphe et par l’extrémité des tentacules qui entraînent et immobilisent les proies par décharge de nématocystes par les cnidoblastes (appareil venimeux représenté par une cellule sécrétrice et sensorielle, voir aussi cet article : agression-du-corail).
La capture est assurée par le courant inhalant du siphonoglyphe et par l’extrémité des tentacules qui entraînent et immobilisent les proies par décharge de nématocystes par les cnidoblastes (appareil venimeux représenté par une cellule sécrétrice et sensorielle, voir aussi cet article : agression-du-corail).Les aliments subissent à ce niveau une première digestion extra-cellulaire grâce à la sécrétion glandulaire de protéases par des cellules spéciales : les cellules caliciformes.
La digestion se poursuit au niveau des filaments mésentériques ventraux et latéraux qui possèdent eux aussi un grand nombre de ces cellules.
Il existe une relation entre l’apparition et la croissance du nombre des zooxanthelles, d’une part, et la régression progressive des filaments mésentériques ventraux et latéraux, d’autre part (Pratt, 1905 ; d’après Grasse et al, 1987). Par exemple chez Lobophytum sp, les filaments mésentériques contiennent un grand nombre de cellules glandulaires et seulement quelques zooxanthelles ; chez Sarcophyton sp les filaments mésentériques sont plus petits, les cellules glandulaires moins nombreuses et les zooxanthelles plus abondantes ; chez Silunaria sp les filaments mésentériques sont très petits voir absents, les cellules glandulaires très rares et les zooxanthelles très nombreuses.
Décharge d'un nématocyste :
 Importance des zooxanthelles :
Importance des zooxanthelles :
Les zooxanthelles sont des algues unicellulaires appelées Dinoflagellés appartenant au genre Symbiodinium, à ce jour plus de 80 espèces différentes ont été identifiées et elles ne sont présentes que chez les coraux hermatypiques. Elles présentent un corps ellipsoïdaux ou sphériques de 7 à 14 µm de diamètre et de couleur brune. Les zooxanthelles possèdent un cytoplasme renfermant des vacuoles qui contiennent des réserves (surtout des lipides), des granules pigmentaires et des cristaux d’oxalate de calcium. Elles sont constituées par un grand corps d’assimilation (vacuole propre aux dinoflagellés symbiotiques), de nombreux chloroplastes situés à la périphérie, un pyrénoïde pédicellé relié à la face interne du chloroplaste par un court pédoncule (il contient la ribulose, enzyme responsable de la photosynthèse) et enfin le noyau.
Photo : zooxanthelles Les zooxanthelles sont localisées au niveau de l’endoderme en position intracellulaire à raison d’une cellule végétale par cellule animale, cependant on a isolé chez certains coraux plusieurs zooxanthelles dans une seule cellule. Elles sont contenues dans une vésicule délimitée par une membrane de la cellule animale : la vésicule périsymbiotique. Elles sont plus abondantes au niveau des parties superficielles de l’animal : disque oral, cœnosarque et tentacules.
Les zooxanthelles sont localisées au niveau de l’endoderme en position intracellulaire à raison d’une cellule végétale par cellule animale, cependant on a isolé chez certains coraux plusieurs zooxanthelles dans une seule cellule. Elles sont contenues dans une vésicule délimitée par une membrane de la cellule animale : la vésicule périsymbiotique. Elles sont plus abondantes au niveau des parties superficielles de l’animal : disque oral, cœnosarque et tentacules.
Selon les auteurs il y aurait entre 1.000.000 et 5.000.000 algues par cm2.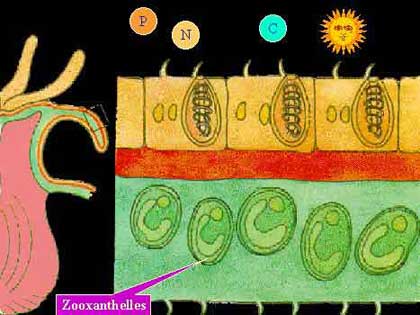 La densité des zooxanthelles et leur répartition dans les tissus sont variables, chez les colonies branchues elles sont moins abondantes vers l’extrémité des rameaux et chez les colonies lamellaires, ce sont les parties supérieures qui renferment le plus de zooxanthelles, les polypes en expansion le jour renferment d’avantage de zooxanthelles, notamment au niveau des tentacules. Cependant la lumière semble être le facteur le plus important, plus la colonie est soumise à l’intensité des radiations et de la proportion en UV plus la densité en zooxanthelles diminue (Stimson, 1997). Pour terminer, le nombre de symbiotes dépend également des besoins nutritifs et du métabolisme du polype, moins il est actif pour capturer des proies plus la densité en zooxanthelles est importante (Grasse et al, 1987).
La densité des zooxanthelles et leur répartition dans les tissus sont variables, chez les colonies branchues elles sont moins abondantes vers l’extrémité des rameaux et chez les colonies lamellaires, ce sont les parties supérieures qui renferment le plus de zooxanthelles, les polypes en expansion le jour renferment d’avantage de zooxanthelles, notamment au niveau des tentacules. Cependant la lumière semble être le facteur le plus important, plus la colonie est soumise à l’intensité des radiations et de la proportion en UV plus la densité en zooxanthelles diminue (Stimson, 1997). Pour terminer, le nombre de symbiotes dépend également des besoins nutritifs et du métabolisme du polype, moins il est actif pour capturer des proies plus la densité en zooxanthelles est importante (Grasse et al, 1987).
Il existe différents pigments photosynthétiques :
- Chlorophylle a
- Chlorophylle b (rare)
- Chlorophylle c
- Caroténoïdes
- Phycobilines
Les caroténoïdes et les phycobilines jouent un rôle fondamental en permettant d’élargir le spectre des longueurs d’onde utilisables (Lesage et al, 2005).
La présence de plusieurs pigments différents permet aux zooxanthelles de s’adapter aux différentes conditions d’éclairement, aussi bien en terme de quantité que de qualité. Cela permet d’optimiser la captation d’énergie lumineuse quelle que soit la profondeur des colonies et les variations d’ensoleillement.
Tous ces pigments présentent un pic d’absorption maximale situé entre 408 et 705 nm, c’est à dire dans la bande bleue du spectre visible (Sprung et Delbeeck, 1996).Grâce aux pigments qu’elles renferment, les zooxanthelles pratiquent la photosynthèse.
Les zooxanthelles utilisent l’énergie lumineuse pour réaliser la photosynthèse. Elle se déroule en deux phases : la première est appelée phase claire et correspond à la transformation de l’énergie solaire en énergie chimique, la seconde est nommée phase obscure (car ne nécessitant pas de lumière) et permet de transformer le CO2 en glucide grâce à l’énergie produite lors de la première phase.
Le taux de photosynthèse dépend de l’intensité lumineuse, il croit avec cette dernière jusqu’à une valeur maximum au delà de laquelle il n’augmente pas et peut même diminuer (photoinhibition) (Schmitz et Kremer, 1977 ; d’après Grasse et al, 1987).
Les zooxanthelles consomment du gaz carbonique et synthétisent au contact du tissu corallien différents produits qui sont pour l’essentiel : du glycérol, du glucose, des acides aminés (essentiellement de l’alanine), et quelques autres acides organiques.
La symbiose est une association qui dure pendant au moins une partie du cycle biologique, entre deux ou plusieurs organismes spécifiquement distincts. Cette association conduit à la formation d’une nouvelle entité biologique : le symbiocosme.
On appel le plus grand des partenaires, l’hôte, et le plus petit, le symbiote.
Sur le plan physiologique, elle permet généralement une meilleure adaptation au milieu ; sur le plan génétique, elle peut être un moyen sophistiqué d’acquérir de nouveaux gènes par transfert latéral.
Lors de la reproduction asexuée, les zooxanthelles sont transmises via les tissus de la colonie mère.
Lors de la reproduction sexuée, les zooxanthelles sont ingérées en pleine eau par les stades larvaires.
Par leur activité photosynthétique les zooxanthelles libèrent de l’oxygène qui est utilisé par le tissu animal, ils jouent donc un rôle important dans le métabolisme du corail.
Le corail utilise certaines molécules produites par les zooxanthelles pour réaliser son métabolisme.
Les molécules organiques fabriquées par les zooxanthelles passent à travers la membrane de l’algue, qui leur est perméable, pour rejoindre le cytoplasme des cellules du polype. Des études ont permis d’identifier le passage à travers la membrane de glycogène et de lipides sous forme de granules (Patton et al, 1977 ; d’après Grasse et al, 1987).
Les zooxanthelles fabriqueraient aussi des vitamines et des hormones utilisées par le polype (Crossland et Barnes, 1977 ; d’après Grasse et al, 1987).
Elles jouent également un rôle dans la fabrication de l’ADN du polype à partir de la thymidine.
Les zooxanthelles peuvent fournir jusqu’à 98% des produits de leur photosynthèse à leur hôte. Elles permettent aussi l’élimination des déchets azotés et phosphorés du corail.
Photo : cytoplasme
 En contrepartie, le métabolisme du polype produit des déchets qui sont utilisés en partie par les zooxanthelles.
En contrepartie, le métabolisme du polype produit des déchets qui sont utilisés en partie par les zooxanthelles.Les produits qui transitent du polype vers les zooxanthelles sont les suivants :
- le gaz carbonique
- les composés phosphorés : les algues symbiotiques utilisent le phosphate produit par le polype afin de réaliser leur métabolisme protidique.
- les composés azotés
- l’acétate : c’est un produit du catabolisme de la cellule animale qui, à l’intérieur des zooxanthelles, est activé puis converti en acides gras dans les chloroplastes (Benson, 1984 ; d’après Sprung et Delbeeck, 1996).
Il faut également noter que le corail fournit une protection, au sein de ses cellules, aux zooxanthelles ; il leur permet ainsi d’éviter la prédation et de lutter contre l’action des sédiments.
Photo : chloroplaste
 Bibliographie et origine des textes : Thèse Vétérinaire sur la physiopathologie des coraux (mai 2007, Julien Vimal)
Bibliographie et origine des textes : Thèse Vétérinaire sur la physiopathologie des coraux (mai 2007, Julien Vimal)NDLR : Chapeau bas l'artiste.
Liens :
http://nanozine.blogspot.com/2006/08/coraux-et-photosynthse-vus-sous-un.html
http://aquariocatss.free.fr/aquariophilie/Recifal-marin/Zooxanthelles.html
www.vet-lyon.fr/bib/fondoc/th_sout/dl.php?file=2007lyon025.pdf
http://nanozine.blogspot.com/2006/10/agression-du-corail-ou-comment-le.html
http://vieoceane.free.fr/paf/ficheb3.html
1 commentaire:
Bel article. Il faut cependant arrêter d'utiliser hermatypique (qui signifie constructeur de récif) comme un synonyme de symbiotique ou de zooxanthellé. Cette erreur, que l'on relève également dans un numéro précédent du magazine, vous conduit à écrire que les zooxanthelles ne sont présentes que dans les coraux hermatypiques. La plupart des coraux mous ont des zooxanthelles et ils ne sont pourtant pas hermatypique.
Noctiluca
Enregistrer un commentaire